Faithfully conveying values and reproducing historical splendor, selected from "Ancient Vertebrates and Ancient Humanities", 1983
Keqin Gao (Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences)
Keywords Shanxi Liulin Late Permian Sawtoothosaurus morphological classification
Executive Summary
This article describes a new genus of copper-toothed dinosaur fossils collected from Liulin, Shanxi, Hmangheaurn lirnlinensit, gen.et sp. nov.)。 According to the comparison of its morphological characteristics with similar fossils at home and abroad, it is believed to be at the same evolutionary level as the genus Seutosanrus. Thus, the fossil's production level can be roughly compared with the Late Permian IV Belt B layer in the Soviet Union.
Serratosaurs are an early reptile group that lived in the Late Permian, and fossils are mostly found in South Africa and the Soviet Union. There have also been sporadic discoveries in the United Kingdom, East Germany, Italy and Tanzania. Domestic reports of such fossils were first reported in 1963, and since then, they have been found in Shanxi and Henan. In the summer of 1980, together with Comrade Yan Huanzhong of the Shanxi District Survey Team, an incomplete skeleton of a serrated dragon was collected from the top stratum of the Shi Qianfeng Formation near Xuecun in Liulin County, including materials such as the right mandible, spine, ribs, shoulder straps, left forelimbs and nail plates. This batch of fossils is described below.
I. Specimen Description
Cup Dragons Cotylosauria Cope, 1894
Procolophonia Romer, propitiated lizard suborder, 1966
Pareiasauridae Cope, 1896
Yellow River Dragon, new genus Huanghesaurus gen.nov.
Characteristics of the genus (see genus species) Huanghesaurus liulinensis.
Yellowhesaurus liulinensis, ep.nov. (Inset 1-7; Plate 1, II)
Orthotype Specimen An incomplete skeleton, including well-preserved right mandibular and part of the left mandible; 13 spines; Left shoulder blade, anterior beak, and beak; Left and right clavicle and interclavian; Left forelimb humerus, ulna, radius, and part of the forefoot bones. In addition, there are several ribs and nail plates. Institute of Vertebrate Paleontology and Paleoanthropology specimen number; V 6722。 Origin and stratification: Xue Village, Liulin, Shanxi; Top of the Upper Permian Stone Thousand Peaks Formation, purple-red sandy mudstone.
Characteristics of species Individuals are large. The mandibular arch is broadly U-shaped. The tooth bone joint is high, and the splint joint is firm. The postarticular process is more developed. The number of teeth is large and compact. The crown of the tooth is elongated vertically and slightly overlapped. The teeth are numerous and evenly distributed. The spine is deeply double concave and the dorsal lateral concave is deeper. The nerve spines are high and thick. The scapula is long, cleithrum groovc .) The acromion is more developed, moderately valgus, and the position is low. The scapula is long, located posteriorly, and its scapula is longer than the black beak part. The beak plate is low and long, and the anterior beak bone is significantly longer than the black beak bone. The collarbone and interclavicle are very thick. The humerus is short and thick, with a large dilation at the proximal end and a very small distal dilation, with strong torsion between the two ends. Both the internal and external condyle holes are more developed. The ulna and radius are long, and the ulna-elbow process (olecranon) is developed.
Specimen Description The right mandible, spine, shoulder strap and left forelimb of this specimen are well preserved, especially the tips of the mandibular teeth, the joint condyles, joint fossa and condyle holes of the shoulder peak, shoulder mortar and forelimb bones, and other identifiable structural features are relatively clear, which is not available to other specimens of the same kind in China. 1. Jaw
The mandibular protrusion of serrated dragons is characterized by the development of a relatively stout protubera-nce at the ventral horacic area. Our specimen is well preserved in the right lower jaw, and although the corner bone is missing, it can be supplemented by the left side. Only the umbras and splint bones are preserved in the lower left forehead.
The dental bone is relatively thick, accounting for about the first half of the lateral jaw. The lateral side of the bone is smooth, and three small holes that may be associated with bone nourishment develop near the joint site. The size and arrangement of these holes is similar to that of the Soviet head ankylosaurus. Since the tooth bone is separated from the splint bone, we can directly observe the structural characteristics of the interior of the mandibular: above the mecklian canal, the tooth bone is significantly thickened, forming the top wall of the myx tube.
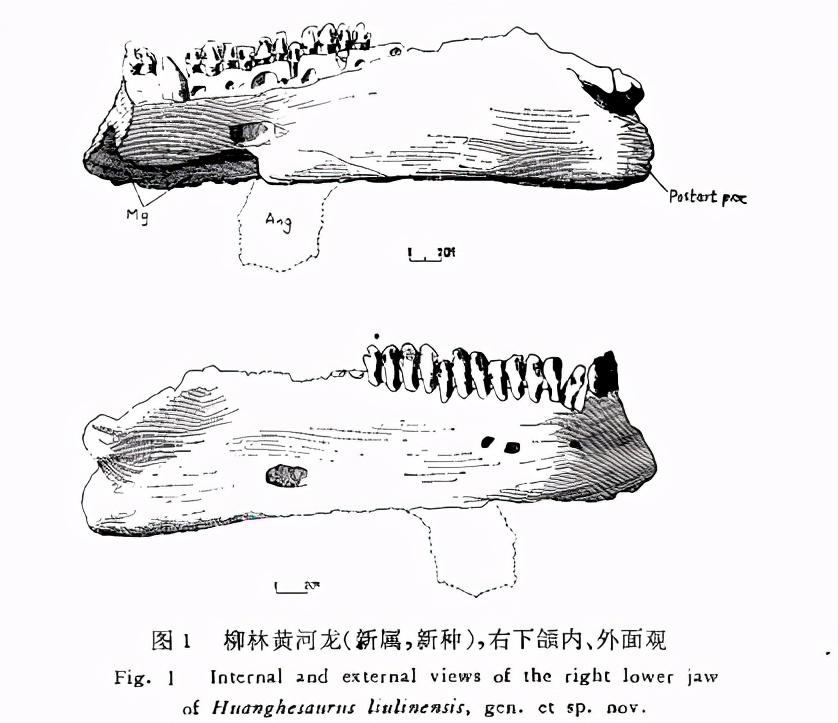
The splint bone is thinner and more exposed on the lateral side of the lower jaw. The bone became unusually thick at the joint of the lower jaw, and the left and right branches healed firmly, and there were almost no traces of the boundary. The joint jaw is about 110 mm high, of which the splint joint accounts for about 55 mm.
The umbra is small, extends very short backwards, and the main body forms a ventral boom of the mandibular. The protrusion is rectangular, medium in size, smoother and more advanced than other types such as Pareiasaurus.
Other bones of the lower jaw, such as the coronal bone, joint bone, anterior joint bone, and superior bone, have all been healed together, making it difficult to distinguish the bone boundaries, but in general, the basic characteristics of the jaws of the serrated dragons are still clearly reflected.
Laterally, the upper bone has a larger opening and appears to be anterior to that of a cephalopod. The medial side of the two articular condyles of the joint bone is larger and slightly anterior; The outer side is small and slightly behind. Between the two condyles is a deep groove. Among the known materials of the same kind, with the exception of the Soviet cephalosaurus, the posterior protrusion of the mandibular joint is generally weak. The V6722 specimen has a more developed posterior joint process and more pronounced muscle marks, indicating that the mandibular augmentation muscle attached to this process is well developed.
The right mandibular tooth bone of the mandibular tooth system has 19 teeth preserved (the last 5 are preserved only the root), plus the total number of unsaved first third teeth should be 20. Among them, the first fourth tooth is not normally developed, the inside and outside are reversed, and the loss of the third tooth may be caused by trauma.
The teeth are compactly arranged, slightly overlapping between the crowns, and the roots are deeply embedded in the bone. The crown of the tooth is elongated vertically, and the lateral convex surface has a more obvious vertical edge extending to the root of the tooth. The medial sides are slightly concave, the middle is raised, and the small tip of the tumor common in other genera is almost invisible on the transverse band of the crown (cingulum). As with the outside, there are vertical ridges that extend from the tip of the tooth to the root of the tooth. A single tooth has up to 17 tips, evenly distributed over the edges of the crown. The total length of this series of mandibular teeth is approximately 220 mm. 2. Spine and ribs
There are 4 cervical and 9 dorsal vertebrae preserved, two of which are well preserved and the other two have broken nerve spines or arthroplasts. 5 of the dorsal pushes are better, the remaining 4 are more severely damaged, and some are only partially preserved.
Cervical spine Serratosaurs have 6 cervical vertebrae, and the 4 connected cervical vertebrae in our specimen represent the last 4 including the pivot. These cervical spines have the following characteristics: large neural tube diameter. The vertebral body is double concave deep and non-lateral, with a more pronounced medioventral ridge and two transverse processes that both develop on the vertebral body – the upper parapophysis and parapophyis.

The pivot is short, the upper and secondary processes are very low, and the spacing between the two is extremely small. The articular surface of the anterior articular process is oblong and slightly inclined toward the midline. The two articular processings extend parallel to the front and extend very short. Both posterior articular processes are not preserved. Specialized nerve spines are not well preserved, but can be seen to be plate-like. The vertebrae are approximately 90 mm high, 95 mm long and 80 mm wide.
The characteristics of the fourth to sixth cervical vertebrae are roughly the same; The anterior and posterior joints protrude obliquely forward and backward, and the lateral extension is small. The nerve spines are slender and slightly thicker upwards, and the apical expansion has two posterior protrusions for anterior axial muscle attachment. The four contiguous cervical vertebrae that have been preserved are about 350 mm in length, and with the addition of the unsaved atlantoaxial and atlantoaxial vertebrae, the specimen is estimated to be more than 450 mm long.
Dorsal vertebrae Because the 9 dorsal vertebrae are scattered and preserved, and the difference between the dorsal vertebrae is far less significant than that of the cervical vertebrae, it is not easy to determine the exact position of each vertebra, but it can be roughly arranged in the order of their front and back.
The vertebral bodies are deeply double concave, and the lateral concaves are also deeper. Several anterior dorsal difficulties, the ventral mid-ridge has disappeared, the parapredic has healed with the upper paradochet and is elevated to the position of the vertebral arch, but there is still a thick ridge connected to the vertebral body. Both the hypovertebral fossa (hypantrum) and the hyposphene are more pronounced. The anterior and posterior articular process protrudes obliquely to the sides and is more wide. The upper paragenesis has not yet fully healed with the anterior articular process. The nerve spines are slenderer than those in the dorsal spines behind them, and the apex is similar to that of the cervical spine, with two protrusions to which the axial muscles are attached
The lateral fovea of the central dorsal vertebra is exacerbated, and there is a well-developed oblique triangular surface in front of it, which may be the location of the interflector. The anterior and posterior joint process extends straight to the sides, with a larger extension and a flat joint surface. The upper parapredicle is completely healed with the anterior articular process, and the subvertebral process and subvertebral fossa are less pronounced than those of the anterior dorsal spine. Below is a measurement of the middle dorsal spine, which helps us understand the general characteristics of the dorsal spine of this specimen.
Full spine height - 300 mm; Vertebral length——— 80 mm;
Nerve spines ——— 100 mm high; Push body width - 50 mm;
Joint process elongation width - 260 mm; The height of the vertebral body ——— 100 mm.
The dorsal vertebrae of the serrated dragons are generally about 15, and the above 9, judging from their structural characteristics, may represent the anterior and central dorsal pushes, while the posterior dorsal, recommended and posterior vertebrae are not preserved.
Ribs Most of the more than a dozen ribs preserved are incomplete, but the neck ribs and dorsal ribs can be distinguished according to morphological characteristics. The neck ribs are short and straight, and the ribs are double-headed. The capicula and tuberculum are developed, and the two are separated in a "Y" shape. The total length measured from the last rib of the left neck is approximately 170 mm. The ribs of the anterior dorsal spine are slender, the small head of the ribs is far from the rib nodes, and the notches are extremely shallow, showing the transitional characteristics from the double head and neck ribs to the typical full head ribs of the dorsal spine. The middle dorsal bone has completely become a full head rib, the ribs are flat and wide and curved inward, and the rib joint surface can be up to 120 mm long. Deeply developed grooves and well-developed pterygic protrusions at the posterior edge of the ribs for intercostal vessels, nerve enrichment, and attachment of the extra-abdominal oblique, spinous, and intercostal muscles. 3. Shoulder straps
The shoulder straps of the V6722 specimen are partially preserved, and all the bones are intact except for the missing right shoulder blade, the black beak bone, and the anterior dark beak bone. Materials include the left shoulder blade, the black beak, the anterior beak, the left and right collarbones, and the interclavian.
The scapula is long, narrow, straight, and slightly sloping backwards. The narrowest above the proximal shoulder is 110 mm. The distal end has a paddle-like spread section with a maximum width of 200 mm. The bone is thicker , about 750 mm long , making it the longest known fossil of its kind. The dorsal anterior margin of this bone is smooth and without a key groove. The shoulders are well developed, rectangular, moderately eloped, and low in position. The supraglenoid fossa is markedly degraded, the pit of the supraglenoid fossa has nearly disappeared, while the supraglenoid buttress is well developed. The medial side of the bone is relatively smooth and the sub-scapular fossa is shallow. Precoracoid
Foramen) has a large, nearly triangular, medial opening at the lower medial end of the scapula.
The black beak plate is preserved only on the left side of the shoulder blade healing, and consists of the anterior beak bone and the black beak bone. The two bones have also healed each other, but the boundary between the two can still be separated from the ridge at the place of healing. The anterior beak is low, significantly longer than the beak, but thinner than the latter. The anterior beak hole is large, round, 35 mm in diameter, and is located anteriorly and below the scapula.
The beak bone is short and thick, and its length is only about two-thirds of the length of the anterior beak bone. The maximum front and rear length of the entire black beak plate is 390 mm. The scapula consists of the scapula and the black beak, and its scapula part is longer than the black beak part. The molars are deep, elongated in a downward posterior direction, about 110 mm wide, and have a maximum length of 230 mm.
The collarbone is thick and well developed, and the width and narrowness of the two ends vary greatly. The ventral lower end is the widest, 105 mm, narrows to the dorsal direction, and is only 55 mm wide at the apex. There are many small neoplasts at the dorsal apex, which are the attachment points of most of the trapczium. The posterior mid-to-lower border of the bone, with thickly developed pterygias associated with the interclavian nodes. The left and right clavicles have a straight length of 510 mm.
The interclavian "T" shape is thick and cumbersome, with an ecros-bar at the front extending about 380 mm to the left and right, and a stem at the rear that is short and wide, with a maximum width of 140 mm. The total length from the leading edge of the transverse band to the rear end of the vertical shank is approximately 320 mm.
4. Forelimbs
Materials include the left humerus, ulna, radius and part of the left forefoot bone, of which the ulna and radius are best preserved, the distal humerus is damaged, but the inner and outer condyle holes are still preserved.
The humerus is short and bulky, with a preservation length of 380 mm, and the estimated full length should be between 420 and 450 mm. The proximal end of the bone is strongly dilated, with a maximum width of 360 mm; The distal expansion is small, less than 200 mm. The two ends twist each other at an angle of about 50 degrees. The backbone is extremely short, columnar, with a cross-section of 55×75 mm.
The proximal anterior dorso-ventral line is thick. Dorsal-pectralcrest is medium in length, about 160 mm. The lateral-medial line is a sleek convex that extends from the proximal articular condyle to the expanded area distal to the diaphysis. Proximal articular surface band, up to 82 mm wide, dichotomy is not pronounced.
The trochlear fossa is deeper. The lateral epicondylar is relatively large, and the supinator process is well developed. The lateral condyle is round and approximately 12 mm in diameter. The entepicondylar is not too stout, but the epicondylar is well developed. The hole is 40 mm long and is formed by a thin outer hole wall bridge arch. The foramen is located near the ventral medial front of the humerus.
Ulna 560 mm in total length. The proximal end is swollen and up to 200 mm wide. The proximal articular surface is 150 mm long and 80 mm wide, and the dichotomy is more pronounced. The two sigmoid processes at the proximal anterior front are more developed, and the sigmoid notch between the two protrusions is also deeper. The elbow process is well developed and is about 120 mm long. The backbone is thinner, with thick wing-like protrusions in the middle and lower sections of the shaft. The distal end of the shaft is slightly thicker and the joint surface is flat.
Radius 390 mm in length, swollen at both ends and contracted in the middle. The recesses on both joint surfaces are deeper, with a maximum length of 140 mm and a maximum width of 85 mm for the proximal ends. The maximum length of the distal one is 135 mm and the maximum width is 95 mm. The diameter of the shaft is 50×70 mm.
The wrists of the forefoot serratosaurs generally consisted of eight bones, and the wrist type was 3·0·1·4. V6722 The specimen's left forefoot preserves only the radial carpal bone, while the remaining seven bones are not preserved.
The radial carpal bone is large, with a length of 90 mm between the near and far ends. The proximal joint surface is an oval convex surface with a maximum length of about 160 mm. The joint surface is surrounded by a ridge-like border that controls the range of motion of the radial-wrist joint. The anterior boundary of the axis is more distal than the axis, indicating that the joint is primarily engaged in forward motion.
Two pieces of the metacarpal bone and the phalangeal bone are preserved, and the finger pattern of the specimen cannot be judged due to the lack of material. As far as is known, the fingering of the sawtooth dragon class is generally 2·3·3·3·1 or 2·3·3·3·2.
5. Nail tablets
Along with the fossil skeleton, more than 20 scattered pieces of nail were also found. According to the shape and size, it can be divided into two categories: one is large and thick, and the size is about 60×80 mm. The dorsal mask is a large central tumor with many small tumors on the sides, which may be a nail patch on the shoulder. The other is smaller, generally around 40×50 mm, thinner than the aforementioned one. The appearance is shell-shaped, and the center tumor and radial fold on the back are relatively weak. This type may be a piece of nail on either side of the dorsal midline or midline. It seems that the animal's leather armor is relatively developed.
Second, comparison and discussion
The V6722 specimen described in this article, according to its morphological characteristics, should undoubtedly be classified in the family Serrates of the order Cuposaurus.
To date, nearly 20 fossils belonging to this family have been found around the world. Among them, Elginia, found in Scotland, England, is a special type of small body with protrusions developed on the head. The Hauboldisaurus and Parasaurus of East Germany are still in doubt, the former with a lower stratum and only nail sheet material, which is now classified as Rhipacosauridae (Kuhn, 1970), while the latter should most likely be incorporated into the Sclerosaurws of the pre-prism family (Kuhn, 1969). Pachypes in Italy have only footprint material. These genera are not very much related to the fossil types described in this article and do not need to be compared in detail. The V6722 specimen is primarily compared to similar fossils in South Africa, the Soviet Union, and China.
Compared with the South African genus, the V6722 specimen was significantly distinguished from fossil types such as Anthodon, Parciasuchur, Bradyseurws and Dalichparia in terms of mandibular morphology, number of cusp, crown features, and morphological characteristics of the acromion, scapula, and beak plate. It is more similar to the genus Serratosaurus, but it is also different. From the differences in these features, our specimens have a longer crown, a relatively developed posterior process of the mandibular joint, a rectangular umbel boom, and a large angle of torsion of the humerus.
In addition to the previously discussed genus of the Same genus as South Africa, the Soviet serrated dragons include rhipaeosaurus, Leptoropha, Para-bradysaurus, Proelginia and Scutosaurus. The first two types, which have primitive features such as small individuals, few tips and slender limbs, were established by Chudinov in 1955 in a separate family, Rhipacosauridae. The family may be closely related to the ancestors of the family Sawtooth. Proelginia has been classified in the genus Cephalopoda, while Parabradysaurus has undergone several taxonomic places before being placed in the Estemennosuchidae family of giants. Of these fossils, the closest to the V6722 specimen is Cephalopodaurus. Although the two are very different in terms of the shape of the mandibular boom and the thickness of the bones behind the head, they cannot be classified as the same genus, but their number of teeth is at most 20, the number of tips is 17, the crown of teeth has elongated characteristics, and the posterior process of the mandibular joint is also relatively developed, indicating that the similarity between the V6722 specimen and the cephalopod is far more obvious than that of other similar fossils. In terms of the number of tips and arrangement characteristics. These two fossil types are basically on the same evolutionary level.
There are four genera of comparable fossils in China, the most important of which are Shihtienfenia and Shansisaurus. Although neither genus has yet to find a jaw or teeth, the differences in the bones behind the head are obvious, suggesting that the new specimen is not suitable for any genus of Shi Qianfeng or Shanxi Dragon. The comparison of features between them can be listed on the following page.
The other two genera in China, Honamia and Tsiyuamia, were established by Professor Yang Zhongjian in 1979. Both genera have only dental material, and new specimens are not easy to compare with their other features, but only
The three can also be distinguished in terms of tooth characteristics. The number of jiyuan dragon's tooth tips is very small, only 7, while there are as many as 17 new specimens, which are very different. Although the tip of the tooth of The Henan dragon is more, its crown is narrower, almost the same width as the root of the tooth, which is also significantly different from the new specimen.
From the above comparison, it is not difficult to see that the V6722 specimen collected from the top of the Qianfeng Formation of Liulin Shi in Shanxi is a new type of serrated dragon, and we recommend naming it Huanghesaurus liulinensis, gen.et sp. nov.), in the family Serratidae.
Historically , the number of species found in South Africa was mostly serrated fossils , with nearly 30 species in 11 genera listed by Brink and Haughton (1954). Nevertheless, the proportion of such fossils in South Africa's Late Permian fauna, which is composed mainly of Dicynodonts and 'Theriodonts). At present, in the Late Permian strata of North China, although there are scattered teeth of diplodonts and mammals, the main fossil types are still only sawtooth dragons. Multiple discoveries of such fossils suggest that they may represent a fauna with sawtooths as the main component. The composition characteristics of such fauna are different from those in South Africa.
Soviet Fossils of late Permian serratosaurs are mainly found in the II. and IV fossil belts of the Dvina Series, especially the latter fossils are abundant. The IV zone is divided into two layers, A and B, of which B is the output layer of cephalopods. The specimens of The Yellow River Dragon described in this paper have more obvious differences in the jaw and tooth characteristics of The Cephalopoda, showing the evolutionary level of the two. therefore. The fossil layers described in this article should be roughly equivalent to the B layers of the IV belt of the Dvina system in the Soviet Union.
This article was completed under the guidance of teacher Ye Xiangkui. And Comrade Cheng Zhengwu of the Institute of Geological Sciences, Comrade Li Fenglin of the Beijing Graduate School of Wuhan Institute of Geography, Zhou Mingzhen of the Institute. Comrades Liu Xianting, Zhai Renjie, Li Chuankui, Zheng Jiajian, Dong Zhiming, Li Jinling and other comrades reviewed WenChou and put forward valuable comments. The plate photos were taken by Comrade Du Zhi, washed and printed by Comrade Zhang Jie, and the illustrations in the text were drawn by Comrade Yang Mingwan.
(Received September 17, 1982)
Reference Essay Dedicated
Yang Zhongjian and Ye Xiangkui, 1963: The first discovery of sawtooth dragons in China. Vertebrate Paleontology and Paleoanthropology, 7(3), 195-212. Yang Zhongjian, 1979: A new late Permian fauna in Jiyuan, Henan. Vertebrate Paleontology and Paleoanthropology, 17(2), 99-113. Cheng Zhengwu, 1980: Mesozoic Stratigraphic Paleontology in the Poor Lands of Shaanxi, Gansu, and Ningxia, Volume II; 115—119。
Boonstra.L.D,1932: Pareiasaurian studies. Pt.VIII—The osteology and myology of the locomoter
=pparatus. B.—Fore limb.An. 8.4/r.Mus。 Vol. 28,pt. 4.
——,1934:Pareiasaurian studies. Pt.IX—The eranial osteology.4n.8.4/r.Mu. Vol.31. pt 1.'
——,1984; Pareiasaurian studies.Pt. XI—The vertebral column and ribs. Awn.8.4/r. Muws. Vol. 31,49—66.