傳統上,人們認為在兩栖動物進化成爬行動物之後,一個進化成哺乳動物,另一個進化成鳥類,但這種劃分早已過時。事實上,哺乳動物的祖先,爬行動物和鳥類,在木炭時期晚期分離了3億多年前。在木炭時代的後期,兩隻兩栖動物同時進化出羊膜穿刺術,并開始适應陸地生活。
以前有一個術語叫"哺乳爬行動物(下洞亞崗/單孔子綱)",但現在逐漸被淘汰,統一稱為妖術(beasts),弓輪廓包含所有哺乳動物及其祖先,"關節弓"是指"固定弓"。蝴蝶結是一個非常特殊的部分,很多人會認為它顯示顴骨側面的高度,其實是錯誤的。它實際上是面部眼窩下方的一側。由于頭骨的每一側都有一個較低的孔(一個太陽穴),是以確定了固定的,內建的弓(在一對時間孔下隻有一對拱門,而不是像蜥蜴形輪廓那樣分成兩對駕駛艙)。弓形輪廓的另一個重要特征是四隻腳位于軀幹下方而不是側面。
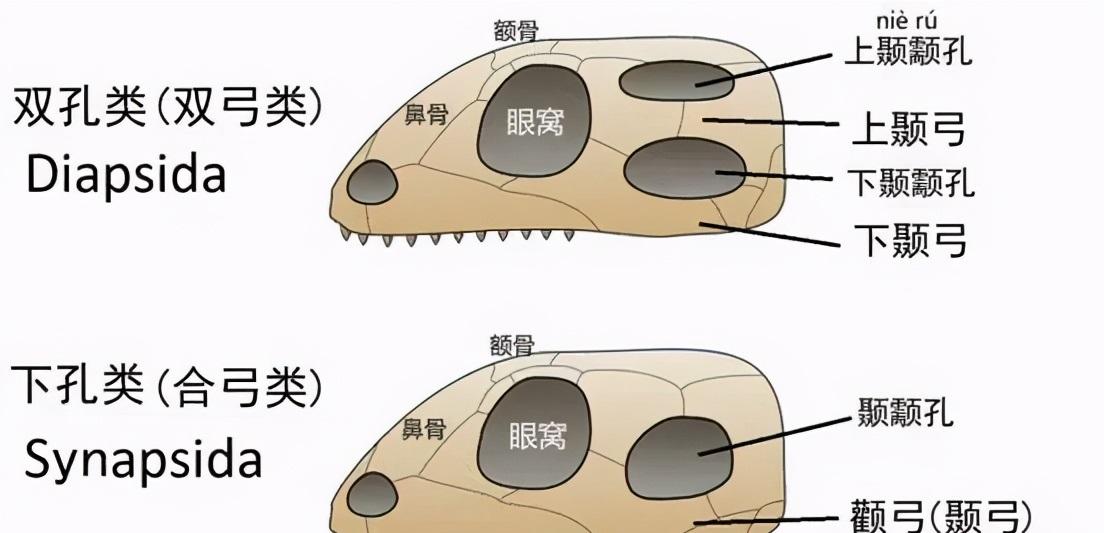
關節弓輪廓和蜥蜴形輪廓的毛孔和拱門

拱門和突出部分
相應地,傳統意義上的鳥類和爬行動物被分為蜥蜴形的輪廓(龍)。這種類型的羊水沒有組合弓,四肢通常在軀幹的側面生長,使其比關節弓更容易直立行走。
巨型節肢動物在3.07億年前木炭雨林崩潰後滅絕,此後幹燥的氣候減少了齒和愛馬仕的優勢。
在古近一代中,聯合弓輪廓在與蜥蜴形輪廓的競争中占了上風。這是由于石器時代晚期,弓形輪廓首先實作了大規模,第一個進化出可彎曲的脊柱,使能力向上移動一個缺口。盤龍武作為最早的一組聯合弓,已經進化出後帆(由長神經棘輪組成的帆),作為後大臣有半圓形的巨大後帆,既有裝甲保護又有一定範圍的溫度調節功能,而且在進化成動物洞後,已經有相當數量的物種進化成恒溫器。
祖先的單弓獸生活在3.06億年前,是已知最早的聯合弓形物種之一,長約50厘米。
生活在木炭時代末期的穿山甲長度超過3米,可能被鱗片覆寫。
基隆,在木炭時代末期開始出現
Hapto野獸始于木炭時代的最後階段,長至少1.5米,以昆蟲和其他小型脊椎動物為食。
生活在2000年代初的楔齒龍長約3.5米,是當時的陸地霸主。
到第二次世界大戰期間,野獸的毛孔達到了前所未有的繁榮和輻射多樣化,并逐漸取代了盤龍。用于野獸洞的孔通常比盤龍的孔大。下颚複雜而強壯。牙齒已經從咬牙的前部特化出來,後部被刺穿,被犬齒撕裂,最後用臼齒咀嚼。而獸洞類也經常抛棄了祖先的盤龍型背帆,畢竟笨重的帆背也是軟肋性質的負擔,一旦折斷就不死不死不傷。
孔隙類的内部進化分支
作為最原始的野獸,巴莫短吻鳄有細長的四肢和眼睛後部的大洞(不像原始弓形輪廓中的其他小孔),是以巴莫短吻鳄可能具有1.5至2米的小咬合力。它們是中等大小的輕型食肉動物,在長方形楔齒龍和更進階的現實世界毛孔之間進化。
俄羅斯的巴莫短吻鳄,生活在兩年一度的中期
此後,恐頭獸在第二世界時期中期占主導地位,所有的恐懼症在咬人時都有上下門牙交錯的特征,大多數恐懼症的頭骨都進化成增厚的頭骨。這些包括最大的食草動物(痂),雜食性野獸(巨獸)和食肉動物(Antio),它們可以長到約4.5米長,重達2公噸,而後者可以超過5米長,但重量僅為500-600公斤左右。在經曆了2000年代中後期在峨眉山的玄武岩噴發(瓜德羅普島時期末期的滅絕事件)之後,恐頭的野獸完全滅絕了,取而代之的是更先進的新野獸。
一頭野獸
大野獸
安提奧野獸
本龍獸屬于一種食草動物,尾巴纖細,體型較小。雙齒獸身材矮小,笨重,桶腰,四肢強壯的大食草獸,頭骨後面的毛孔變大,可以容納較大的下颚肌肉,當嘴巴閉合時,下颚閉合産生強烈的切割作用,使雙齒獸能夠處理堅硬的陸地植物。
龍獸的肢體獸
雙齒獸
到2000年代後期,陂齒獸,獸頭獸和犬齒獸作為進化最快的三種野獸在各大洲蓬勃發展。其中,石斑獸是極其兇猛的捕食者,最初隻是一隻狗大,經過數百萬年進化成一隻4.5米長的巨大食肉狼蜥蜴。獸頭亞眼和犬齒動物亞眼作為真正的獸牙有次颚(在口中分離呼吸和消化道),沒有胫骨,腳趾骨也已淪為類似形式的哺乳動物,其中一些已經進化成真感恒溫動物,其中犬齒動物是現代哺乳動物的祖先。冷血動物和溫血動物之間沒有明确的界限,如箭魚、鮪魚、鼠鲨、棱皮龜,它們是中溫動物,通過燃燒身體化學将溫度調節在一定範圍内,但不能保持絕對恒定的體溫,這是從冷血動物到溫血動物的過渡。進化成恒溫器是不同物種融合和進化下的總趨勢,除了鳥類和哺乳動物之外,爬行動物的Tyga蜥蜴,魚類中的月亮魚是溫血動物,鳄魚祖先在三疊紀時期也是溫血動物但後來退化。盤龍帶背帆其實是一種中溫動物,以獸頭和犬齒動物都有一部分比較先進的物種,已經彙聚成現代意義上的溫血動物。
陂齒獸的狼蜥捕食龍
從2000年代末到三疊紀早期,野獸的頭部是南非的捕食者。
鮑什獸是典型的進階食草動物頭
卡倫人是一種多齒,發達的犬齒獸,可能對小魚或昆蟲具有水生掠食性
野獸頭部的鼻腔Ganosuchus有一個凸起的脊,被認為是第一個進化出恒溫功能的鼻腔。
Dwyna生活在第二世界末期的俄羅斯,是一種小型雜食性犬齒獸,門牙小,犬齒2顆,臼齒10至14顆。
犬齒獸最早出現于第二世界末期,具有完整的次級下颚,進食時仍能呼吸,嘴巴和鼻子寬,四肢直立。
對于盤龍和毛孔,僅從化石中很難判斷它們是否進化出與現代哺乳動物相似的乳房和胎兒功能。
古代蜥蜴形輪廓的情況無法與聯合弓輪廓相提并論,繁榮程度遠低于後者,化石遺骸也不是很豐富,體型也大多很小。
大約3.12億年前的石器時代晚期蜥蜴是蜥蜴形群中已知最早的成員,長約20厘米,牙齒和昆蟲很小。
大鼻龍是一群小型的蜥蜴狀爬行動物,在白垩紀晚期到古新世第二紀幸存下來。
蜥蜴輪廓的内部進化分支
亞爬行動物是最原始的爬行動物,它保留了更原始的特征,例如強壯,低垂的身體和顱骨背面的大胫骨,盡管它在顱骨後部也有兩個開口,但腭裂的程度通常是保守的,是以被認為等同于沒有孔(即, 無穿孔基質)。目前,遺傳分子研究表明,海龜雖然沒有毛孔,但很可能是其雙穴亞群祖先在進化過程中失去了毛孔,龜在近親關系上更接近主要龍種,亞爬行動物現在已經完全滅絕。真正的有毛孔的爬行動物是當今爬行動物的支柱。
根據現有的研究,最初的古代蜥蜴,包括森林蜥蜴,出現在石器時代晚期,雖然頭骨沒有洞,像現代一樣,但它們在親緣關系上更接近真正的爬行動物。現在可以識别的木炭時代晚期的亞爬行動物主要出現在3.03億年前的Erpetonyx和生活在木炭時代末期和第二古新世開始時的龍中期。
Erpetonyx的化石
Erpetonyx假想圖可能是肉食性的,可能具有多種骨骼特征,纖細的身體,大爪子和強壯的肌腱附着點。
中國龍的化石作為闆塊漂移學說的證據
中龍長約1米,細長的嘴适合在鹽湖中釣魚
在雙年展期間,亞爬行動物比真正的爬行動物繁榮得多,前棱柱蜥蜴是副爬行動物的重要進化分支,出現在中馬科動物中,包括各種動物,包括小型,蜥蜴狀的前遲鈍蜥蜴,以及大型,厚裝甲的食草食肉食肉體鋸齒龍。
蘭花短吻鳄長約75厘米,是第二紀元的标志性亞爬行動物。
鋸齒狀的龍有厚厚的盔甲,可以長到3米長,重達600多公斤。
真正的爬行動物在進化過程中分為主龍形的子輪廓和鱗狀的龍形兩個主幹。兩者的差別在于鱗狀龍形牙弓不完整,主龍形頭骨保留完整的雙形洞,鱗狀龍形端牙(喙狀蜥蜴)或側齒(蜥蜴和蛇),主龍形槽牙(包括早鳥);主要的龍形類有上颚孔(前毛孔,即眼窩和頭骨兩側鼻孔之間的一對孔),鱗片龍形物種沒有;鱗狀龍形四肢位于身體外側,運動時脊柱左右擺動,主龍肢靠近身體(恐龍和一些三疊紀主龍肢立在身體正下方,腹部遠離地面),運動時脊柱保持筆直;主龍心室分離度較高,可利用安全氣囊和支氣管雙重呼吸,主龍形心有四腔,鱗龍形類隻有三腔。鱗龍形動物基本上是冷血動物,而主要的龍形動物則有相當比例的溫血動物。主要的龍形類可以了解為蜥蜴形輪廓内的貴族。
二紀元中真正的爬行動物大多還沒有分化到鱗龍和主龍形的程度,更具代表性的是陽蜥蜴、空尾蜥蜴、克勞迪奧蜥蜴等。這一時期鱗龍和主要龍的化石很少見,與蜥蜴形輪廓一般被拱門壓制的情況有關,鱗龍中最原始的蜥蜴出現在第二紀晚期(三疊紀之前缺乏完整的化石), 而主要的龍代表了2.6億年前開始出現的原始龍。作為各種主龍的祖先,原龍體型修長,外觀與蜥蜴相似,脖子、四肢長約2米,其體型表明它們可以快速移動,據推測主要是昆蟲為食。古代鳄魚,曾經被歸類為恐龍祖先和該群體的牙齒,在2000年代後期和三疊紀的長興時期開始時生活在俄羅斯。古鳄魚長約1.5米,四肢很短,向兩側伸展。作為類似鳄魚的動物,它們缺乏真實鳄魚的堅硬鱗片,骨骼在性格上更原始。它們最獨特的特征是上颌前部的前上颚明顯向下突出,并在下颌上方突出。這些主要龍可能生活在動物屍體上,是以這些捕食者在第二次世界大戰結束時逃脫了大規模滅絕,并在三疊紀後期迅速輻射以進化出豐富多樣的主要龍。
楊氏蜥蜴捕食蝸牛
一隻在2000年代後期滑翔的空尾蜥蜴
半海克勞迪奧蜥蜴
雙年蜈夜期晚期的主要俄羅斯龍
原創龍的想象
第二紀元末期的古代俄羅斯鳄魚
總的來說,雖然第二王朝時期的兩栖動物在木炭時期的水域中仍延續着大規模的失齒兩栖動物,但弓也是二疊時期的主要統治者之一,拱門引領了兩重時期的主要生态遺址,唯一的缺點就是未能進入海洋。而蜥蜴形的輪廓一般隻能縮小在一些偏僻的角落,在化石形成時留下的存在感不是很強。