目前,大多數生态學研究都基于物種水準,将物種作為應對氣候變化的單一,非分化的實體。有研究表明,物種水準的生态位是保守的,有研究表明物種的生态位已經分化。物種在分化時間,進化獨立性和遺傳特異性方面是不同進化線的集合。對于系統地理結構分布廣泛的物種,在物種水準上應用綜合譜系的生态位模型(ENMs),很難準确描述适應不同氣候條件的不同遺傳子系統之間生态位的進化動态。此外,需要觀察和研究物種生态位動态下不同分支的形态演變和現象質可塑性。
青藏高原由于印度洋闆塊與亞洲闆塊的碰撞而繼續向北上升,在青藏高原周邊山區形成錯綜複雜的山谷和劇烈的氣候波動,為亞種分化和生态位的進化提供了多樣化的生态機遇。藏牙突分布于青藏高原東南邊緣,橫跨喜馬拉雅山、青藏高原、橫貫山脈和廬山山脈,分布在海拔2400m~5270m,分布廣泛,為研究所學生态位演化和亞型間形态演化提供了良好的研究材料。
由聯合首都師範大學上司的中國科學院成都生物研究所兩栖爬行動物多樣性與保護項目組的研究人員蔣建平和謝峰等人利用生态位模型(生态位模型,ENMs)和分類(環境主成分)基于多年采樣的生物資料進行分析, PCA-env)、系統-發生比較法(系統發育比較法,PCMs)和系統發生最小乘法(系統發育一般最小二乘法,PGLS)探讨了藏牙突分支之間生态位分化水準和生态分化的具體形式。對生态分化貢獻最大的氣候因素和形态進化反應。
結果表明,線粒體基因組顯著分化為6個分支,其中3個(分支E.A、E.B和E.C)發生了顯著的生态分化(基于ENMs和PCA-env方法:Schoener的D小于預期的Schoener's D值,雙向生态相似性檢驗P<.05,則否定原假設, 即發生生态位分化)。在這種情況下,生态水準的差異化主要是由生态水準的膨脹而不是生态水準的填充引起的(生态位置的擴張:生态水準的轉移與局部生态>U;生态水準的填充:生态水準的轉移與當地的生态位置U>E,見原始表3)。環境主要成分PC3(主要由BIO2和BIO3組成)、PC4和PC5(主要由BIO6和BIO7組成)次元環境因子遵循布朗運動與系統發展的關系,促進生态位的演化,這與促進頭部長期進化的環境因子幾乎一緻。此外,生态位置的擴張引起的生态位分化确實伴随着形态進化(例如,具有強系統發育信号的頭部長度(SVL))和表型可塑性(例如前臂和手長(LAHL),腿長(HLL)和腳長(FL)。本研究為廣泛分布物種各分支的生态位置演化和形态特征變化提供了研究模型和理論架構。
這項研究的結果名為Anura:Megophryidae,發表在Ecoology and Evolution上。該研究工作由青藏高原第二次綜合科學考察、生态環境部生物多樣性調查評估項目、四川省科技廳基礎條件平台建設和中國生物多樣性觀測網資助。
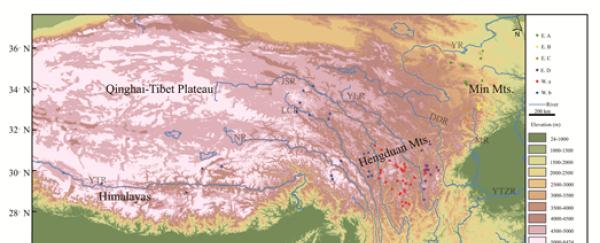
圖 1.發生每個分支的站點分布

圖 2.(a) 家譜關系的地理範圍及其基于最小凸邊法的擷取;(二)各分行的适當分布區域;(c)沿着氣候(PC)和海拔(海拔)的主要組成部分重建祖先關系。
圖 3.(a) 環境變量的三維聚合分析;(b) 沿氣候變數BIO3的每個分支的生态分布;(c) PCA-env分析中涉及的主要成分的因素和貢獻的相關性;和(d-e)生态位等效測試結果;(f-g) 生态相似性的雙向測試結果;(h-i)生态動力學分析結果,紅色表示生态站點的擴充,綠色表示穩定的生态站點,藍色表示生态站點的擴充和穩定的生态站點重疊區域
資料來源:中國科學院成都生物研究所