目前,大多数生态学研究都基于物种水平,将物种作为应对气候变化的单一,非分化的实体。有研究表明,物种水平的生态位是保守的,有研究表明物种的生态位已经分化。物种在分化时间,进化独立性和遗传特异性方面是不同进化线的集合。对于系统地理结构分布广泛的物种,在物种水平上应用综合谱系的生态位模型(ENMs),很难准确描述适应不同气候条件的不同遗传子系统之间生态位的进化动态。此外,需要观察和研究物种生态位动态下不同分支的形态演变和现象质可塑性。
青藏高原由于印度洋板块与亚洲板块的碰撞而继续向北上升,在青藏高原周边山区形成错综复杂的山谷和剧烈的气候波动,为亚种分化和生态位的进化提供了多样化的生态机遇。藏牙突分布于青藏高原东南边缘,横跨喜马拉雅山、青藏高原、横贯山脉和庐山山脉,分布在海拔2400m~5270m,分布广泛,为研究生态位演化和亚型间形态演化提供了良好的研究材料。
由联合首都师范大学领导的中国科学院成都生物研究所两栖爬行动物多样性与保护项目组的研究人员蒋建平和谢峰等人利用生态位模型(生态位模型,ENMs)和分类(环境主成分)基于多年采样的生物数据进行分析, PCA-env)、系统-发生比较法(系统发育比较法,PCMs)和系统发生最小乘法(系统发育一般最小二乘法,PGLS)探讨了藏牙突分支之间生态位分化水平和生态分化的具体形式。对生态分化贡献最大的气候因素和形态进化反应。
结果表明,线粒体基因组显著分化为6个分支,其中3个(分支E.A、E.B和E.C)发生了显著的生态分化(基于ENMs和PCA-env方法:Schoener的D小于预期的Schoener's D值,双向生态相似性检验P<.05,则否定原假设, 即发生生态位分化)。在这种情况下,生态水平的差异化主要是由生态水平的膨胀而不是生态水平的填充引起的(生态位置的扩张:生态水平的转移与局部生态>U;生态水平的填充:生态水平的转移与当地的生态位置U>E,见原始表3)。环境主要成分PC3(主要由BIO2和BIO3组成)、PC4和PC5(主要由BIO6和BIO7组成)维度环境因子遵循布朗运动与系统发展的关系,促进生态位的演化,这与促进头部长期进化的环境因子几乎一致。此外,生态位置的扩张引起的生态位分化确实伴随着形态进化(例如,具有强系统发育信号的头部长度(SVL))和表型可塑性(例如前臂和手长(LAHL),腿长(HLL)和脚长(FL)。本研究为广泛分布物种各分支的生态位置演化和形态特征变化提供了研究模型和理论框架。
这项研究的结果名为Anura:Megophryidae,发表在Ecoology and Evolution上。该研究工作由青藏高原第二次综合科学考察、生态环境部生物多样性调查评估项目、四川省科技厅基础条件平台建设和中国生物多样性观测网资助。
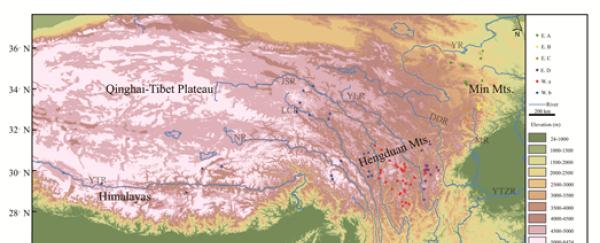
图 1.发生每个分支的站点分布

图 2.(a) 家谱关系的地理范围及其基于最小凸边法的获取;(二)各分行的适当分布区域;(c)沿着气候(PC)和海拔(海拔)的主要组成部分重建祖先关系。
图 3.(a) 环境变量的三维聚合分析;(b) 沿气候变数BIO3的每个分支的生态分布;(c) PCA-env分析中涉及的主要成分的因素和贡献的相关性;和(d-e)生态位等效测试结果;(f-g) 生态相似性的双向测试结果;(h-i)生态动力学分析结果,红色表示生态站点的扩展,绿色表示稳定的生态站点,蓝色表示生态站点的扩展和稳定的生态站点重叠区域
资料来源:中国科学院成都生物研究所