本研究通過便攜式腦電圖(EEG)同時記錄了12名高中學生在正常課堂活動中的大腦活動,并通過一種新穎分析技術--完全互相依賴(total interdependence ,TI)--來評估群體神經一緻性,結果表明學生大腦活動同步程度可以預測學生課堂參與度和社會動态,腦同步可能是由共同注意驅動的動态社互動動神經标記。同時這項研究驗證了在自然環境中研究群體互動神經機制的一種有效方法。文章發表在Current Biology雜志。
【研究問題】
雖然目前對腦如何支援動态群體互動知之甚少,不過各種研究已經開始通過比較多人在各種(半自然)任務中的大腦反應來解決這個問題。這些實驗揭示了刺激特性、個體差異和情境因素如何限制個體神經活動的相似性和差異性。但是,大多數研究都受到各種限制:缺乏參與者直接面對面的互動,通常僅限于二進制組沒有跨時間的社會動态,而且很少研究自然環境下的社會行為。教室提供了實用的、重要的生态自然的環境,同時還提供了一個由教師引導一系列活動的半受控環境使得能夠在整個學期中系統地測量參加了一系列課堂活動(在11個50分鐘課堂中重複進行授課、觀看教學視訊并參加小組讨論)的學生大腦活動和行為。研究假設認為一組學生的同步神經活動可以預測(并支撐)課堂參與和社會動态。那麼當學生感到與材料或彼此聯系互動時,大腦在形式和量化意義上“同步”嗎?為了研究這些問題,打破實驗室壁壘使用低成本便攜式腦電圖(EEG)系統結合一種新穎分析技術研究現實世界中小組互動過程中參與者的神經标記進而來探索個體間大腦活動的同步性:完全互相依賴(TI)(圖1A-1C)。
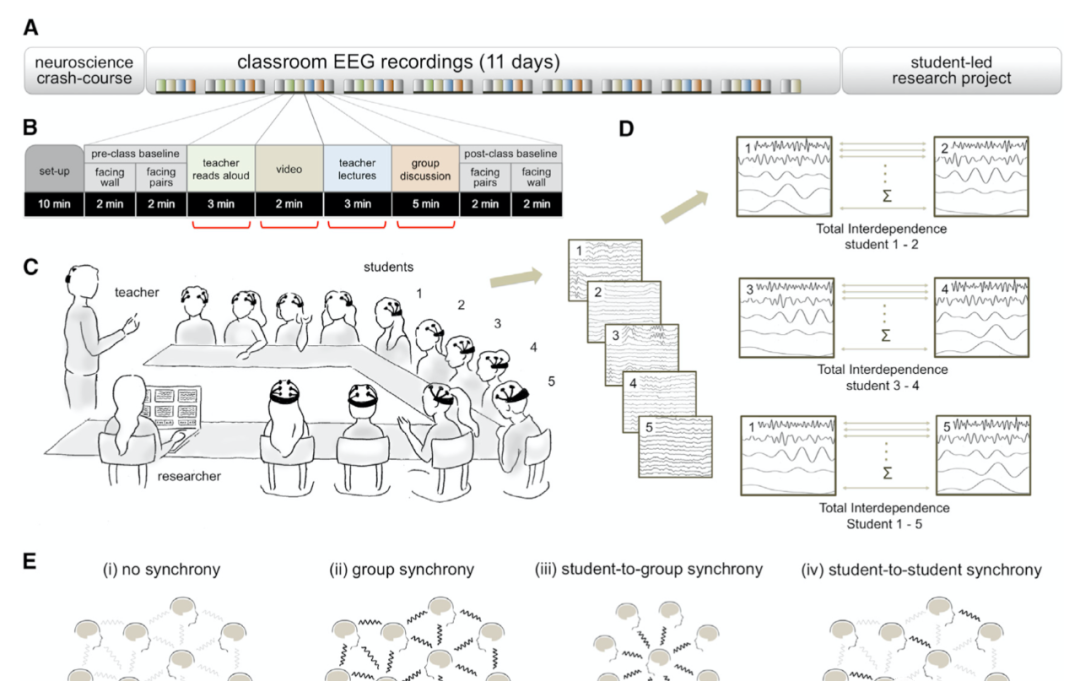
圖1實驗設計、程式和基本原理
A實驗時間表。秋季學期以神經科學速成課程開始,随後3個月内進行11次資料收集。在春季學期中,學生設計并執行自己的原創研究項目。
B實驗過程。在11次EEG記錄中一緻進行視訊、授課和小組讨論等任務。其他任務有選擇出現。計算各教學方式下的平均TI值(紅色标記)。
C佩戴EMOTIV EPOC頭戴式腦電帽的12名學生課堂配置圖。便攜裝置讓學生可以同時作為參與者與實驗者。
D将學生原始EEG信号分解為頻率段(1–20 Hz,0.25 Hz分辨率),并計算每對學生各頻率腦相幹總和進而得出腦同步(TI)。TI量化了整個頻譜的大腦相幹性,進而可以對感興趣的大腦信号進行資料驅動識别。E在多個社會性水準上利用TI分析腦同步性:組同步(對組中所有學生對TI值取平均值)(ii);個體-小組同步(給定學生與其所有同伴TI平均值)(iii);以及學生與學生同步(成對學生TI值)(iv)。
【研究方法】
被試 12名健康無神經系統疾病史17-18歲高中生。剔除兩名女孩資料,頭發體積和頭部形狀導緻資料品質較差。
總程式:
在正式實驗之前對學生進行基礎知識教育訓練以便學生熟悉EEG裝置來降低裝置設定時間。随後記錄他們在11次半正常課堂中的EEG,課程内容為正常生物學課程。每堂課開始前後進行三種不同的基線記錄:面對牆壁(2分鐘;第2-4和8次課程由于時間限制而跳過)、面對小組(第4、6、8和10次課程,2分鐘)或面對面(第5、7、9和11次課程課前2分鐘)。每節課都根據學生位置來确定配對(圖3C),實驗者随機配置設定學生座位。在安裝EEG腦電帽時學生填寫課前問卷。
教學方式:
如圖1所示,課堂活動包括四種主要教學方式:老師給學生朗讀講稿三分鐘;接下來,學生觀看一段與課程主題相關的兩分鐘教學視訊;老師接着授課三分鐘;最後,學生就課堂主題進行五分鐘小組讨論。朗讀因達不到學習目标而在五節課後終止。所有11個課程都包含視訊、授課和讨論,對每個學生每天學習情況和教學方式進行資料分析。
問卷調查:
EEG記錄之前收集學生社會特質資訊,包括群體親和力(見圖2E)和移情指數(圖2F)。此外,學生填寫課前和課後問卷(共9次),詢問學生關注度(圖2F)以及對不同部分的喜愛程度(每日評分;圖2A)。所有課程結束後,再次詢問他們對四種教學方式的總體享受程度(學期後評分;圖2A),對老師的喜歡程度(教師喜歡程度;圖2E),以及對同伴的親密程度(親密程度評級;圖3C-D)。
EEG資料采集:
在11節生物課程(圖1)中記錄12名學生同時發生的腦電活動,學生在EEG記錄中盡量減少明顯運動。14導emotiv EPOC無線EEG腦電帽與OpenFrameworks軟體包配合使用,将12名學生的腦電資料同時記錄到一台計算機上,emotiv EPOC硬體規格見圖S1A,腦電帽見圖S1B,錄音軟體圖形使用者界面截圖見S1E。 電極F7-F8記錄水準眼電,AF4-F8記錄垂直眼電。

圖 S1 A emotiv-EPOC腦電帽硬體規格。
B emotiv-EPOC腦電帽側視圖。
C電極位置俯視圖,分析中包含的位置用綠色标記。
D兩名佩戴EEG腦電帽的示例學生側視圖。同步分析中包括綠色電極,用做h-EOG和v-EOG的電極分别用橙色和灰色标記。
E為本研究開發的錄音軟體的截圖。
F 眼、頭運動僞影回歸分析:F1上下眼動時v-EOG與雙極EEG電極的相關性;F2左右眼動時h-EOG與雙極EEG電極的相關性;F3 回歸前後的方差比;F4 使用模型的不同部分進行回歸前後的方差比。
EEG資料品質判斷:圖S1C為電極位置俯視示意圖,其中包括用于分析的電極(綠色)。盡管電極位置已經被映射到标準10/20布局上,實際電極位置在學生之間不一緻,腦電帽在每個學生頭上的位置略有不同(圖S1D)。對于每對被試,分析機關是為每個電極對(在圖S1C中标記為綠色)計算的TI,然後被試之間的所有可用電極對取平均值(圖S3E)。圖S3A-B顯示了代表性腦電帽14個電極的原始EEG。使用回歸方法去除EEG資料中眼動、頭動(詳細算法及可行性見附S1F)。
通過多個對照實驗,即雙音調ERP實驗、手指移動實驗、其他學校重複實驗,和“完整性檢查”,即對比睜眼(EO)-閉眼(EC;各2分鐘)EEG,來驗證:資料确實可靠和可解釋(圖S1F),并确定完全互相依賴是一種可以跨天、跨群體和學校複制(圖S2F)的有效捕獲共同注意力同步神經活動的方法(詳細算法及可行性見S2A-S2F)。
S1F 使用回歸算法從EEG資料中剔除眼動、頭動影響。
自然實驗條件下,從腦電測量值計算腦同步性時存在一個主要問題:是否由腦電測量中的同步非腦信号驅動。目前實驗背景下,學生參與各種形式的課堂互動時有一些受周圍環境或刺激所引起的非腦源眼睛或頭部運動。例如,跨腦同步性可以由學生同時追蹤重要移動刺激的同步眼動驅動。類似的非大腦相關同步性可能是由環境或刺激物驅動相似肌肉産生壓力或張力引起,進而在EEG記錄中産生相似僞影。為了估計眼、頭運動對腦同步性的影響,從腦電記錄中回歸出四個解釋變量來表示這些僞影,并重新計算TI(完全互相依賴)。
使用emotiv EEG腦電帽内置的兩軸陀螺儀對頭部運動進行評估,并與EEG電極同步采樣。陀螺儀提供兩個變量,?x還有?y,分别表示圍繞?軸和?軸的固定分辨率(1°/?)的角速度。?軸是穿過被試耳朵上方腦電帽底座的“水準軸”(圖S1B)。?軸是沿垂直于腦電帽臂和X軸的方位角方向的“垂直軸”。為了消除腦電資料中頭動,這兩個變量被用作回歸函數。emotiv腦電帽中的陀螺儀具有1°/?的固定分辨率導緻資料階梯式分布而非平滑分布。為了避免在回歸後階梯分布污染EEG資料,對陀螺儀變量進行30Hz低通濾波,進而将階梯式分布轉換成适合于腦電資料回歸計算的平滑連續變量。
眼動通常使用眼電(EOG)進行評估,眼上下方電極監測垂直眼動(v-EOG),左右外眶處電極監測水準眼動(h-EOG)。由于本實驗沒有EOG,是以尋找了兩個電極替代EOG。emotiv電極空間布局(圖S1C-D)明顯表明最接近h-EOG的是電極F7-F8內插補點,其位于水準眼動平行線上且是離眼肌肉最近的電極,是以預計它們的水準眼動敏感度最高垂直眼動敏感度較低。v-EOG最合适的替代電極是AF4-F8和AF3-F7,由于兩組在計算上一緻,是以隻選擇AF4-F8(圖S1F-1)用于後續分析。雖然替代電極的空間位置與實際v-EOG明顯不同,但由于電場随着距離平方的倒數而減小此替代仍能捕捉垂直眼運。也是以,垂直眼動産生的電場在電極AF4和F8中具有相同的極性(都位于眼肌上方)F8值卻比AF4更強,是以其差異可以捕獲垂直眼動産生的電場。由于電極F8和AF4也有水準偏移,它們也會捕捉水準眼動。單一EEG電極替代v-EOG和h-EOG并不可取,因為單一電極攜帶着來自整個大腦的混合電場,而将它們還原出來将消除大量的大腦活動,而雙極替代則靠近眼睛且對來自眼部的強局部電場梯度更加敏感。
為了測試F7-F8和AF4-F8替代EOG是否可行,對一名被試進行了兩次替代實驗,同時記錄EEG(使用emotiv腦電帽)和EOG(使用單獨電極)。在第一個替代實驗中,被試被要求上下移動眼睛一分鐘。在第二個替代實驗中,被試被要求左右移動眼睛一分鐘。真正v-EOG放置在右眼上下方,真正h-EOG放置在眼眶外側。EOG和EEG信号進行1-10Hz帶通濾波,該頻率範圍通常包含眼動僞影的主包絡(由于emotiv有效頻率範圍僅到43Hz,是以無法獲得高γ僞影)。首先,上下眼動過程中計算v-EOG與F7-F8和AF4-F8之間的Pearson相關系數(替代實驗1)。如圖S1F1所示, AF4-F8與真正v-EOG具有更高相關,适合捕捉垂直眼動。類似地,計算左右眼動時實際h-EOG和兩個替代電極的相關性,如圖S1A2所示,兩者具有最高的相關性,這證明了它捕捉水準眼動的能力。v-EOG與AF4-F8相關顯著高于h-EOG與F7-F8相關。這可能由于 AF4-F8對産生強電場的眨眼也很敏感,增加了與實際v-EOG信号的相關性。h-EOG和F7-F8對眨眼的敏感度要低得多。這些結果說明F7-F8和AF4-F8可替代h-EOG和v-EOG用于從腦電資料中回歸出與眼動相關的殘餘僞影。回歸分析中 F7-F8将被稱為?h, AF4-F8将被稱為?v。兩個陀螺儀輸出變量也用作多元線性模型中EEG電極回歸參數。
在下面多元線性方法中,使用主成分建立模型,使用主成分分析(PCA)是因為EEG電極之間、兩個替代眼動間以及陀螺儀輸出變量之間共線度很高。應用PCA去除它們的共線性。相關測量正交分解為不相關分量的相似模組化方法的有效性已經在腦電和腦磁資料分析中得到了證明。線性回歸公式如下:
?pca=?+?∙?+?pca
Ypca是腦電資料主成分。矩陣每一行對應一個主成分,每列對應一個樣本。
?pca =?∙?
?是主成分系數矩陣,它将EEG電極映射到主成分。每一行對應一個主成分,每一列對應一個EEG電極。
?包含兩組回歸變量。?pca包含兩個陀螺儀變量的主要成分Gx還有?y。?pca包含EOG替代電極主成分Ph還有?v。?包含每個腦電電極線性模型常數。?包含每個回歸變量和每個模組化電極的斜率系數。每一行對應一個電極,每一列對應一個回歸變量。
?pca包含無法由給定回歸變量模組化的剩餘EEG資料。這個殘差在主成分空間,每一行對應一個主成分,每一列對應一個樣本。這些殘差資料可以簡單地與主成分系數矩陣的逆相乘,然後再投影到腦電電極空間。
?= ?-1∙?pca ?是去除回歸變量調制後的腦電資料最終殘差,在此基礎上重新計算腦同步性(TI),并與回歸前最初計算矩陣進行比較。
在将上述模型應用之前對EEG資料進行0.5Hz高通濾波并去除線性噪音,陀螺儀資料經過30Hz低通濾波(用于将其階梯式波動轉換為連續波動)并去除線性噪音。在進行PCA和多元模組化之前,剔除所有噪音段(與原始TI計算所用的資料完全相同)。多元模型由使用協方差權重(“cwls”)最小二乘法的MATLAB “MVRecurse”程式計算。盡管最初采用了極大似然法,但如Royston多元正态性檢驗所示,模型殘差的多元分布不是正态分布,是以最終采用了最小二乘法。
在11次不同實驗期間将模型應用于每個記錄資料。對總共90個腦電資料進行了多元模型估計,并對眼睛和頭部運動進行了回歸,留下了一個殘差資料集。為了評估回歸對原始資料的影響,估計了90個資料和每個電極回歸前後的方差比。圖S1F3顯示了每個電極的平均方差比值。灰色陰影表示所有使用資料的标準誤。方差降低幅度最大的三個電極是EOG替代電極F8、F7和AF4。F8是兩個回歸函數的組成部分,其方差減少率最高。AF3方差降低沒有AF4的方差降低那麼多,可能是因為垂直代理變量?v不僅捕捉到垂直眼動,還捕捉到頭部左側對稱位置不存在的大腦右側局部活動(與眼動預期相反)。橫向右側電極F4、FC6、T8和P8的方差減少高于其對稱左側電極(F3、FC5、T7和P7),不過左側電極也有10.3%-16.5%的顯著方差減少。方差減少呈現出從前到後和從外側到内側的下降,枕葉和頂葉電極方差減少最低。
回歸對資料的影響的最終評估方法是檢查回歸變量代表的方差減少多大程度上能歸因于眼睛和頭部運動,這通過省略表示頭部運動的陀螺儀回歸變量模型系數來實作。在此基礎上,計算該模型殘差和方差縮減率。這個比率代表了原始腦電資料中存在多少眼動僞影。然後重複同樣的程式,但省略眼動回歸系數,保持頭部運動回歸系數,這表明原始腦電資料中存在多少頭部運動僞影。如圖S1F4所示,大部分方差減少歸因于眼動回歸(藍線)。陀螺儀回歸系數(紅色)的方差降低仍然很低,低于5%,并且在電極中均勻分布。這表明,實驗過程中頭部運動是有限,EEG資料沒有受到諸如頸部肌肉和電極運動僞影等伴随僞影的污染。在同一圖上,再次顯示完整模型的方差比(黑線),以便進行比較,如預期的那樣,該方差比等于1減去兩個簡化模型的方差比之和。
總之,本節表明:資料中的眼動僞影可以用額葉電極替代EOG來捕捉;從EEG資料中回歸出眼、頭部運動僞影後,所有電極殘差顯著降低,枕、頂葉區域的殘差方差最小;方差的減少大部分是由于眼動回歸系數。(這表明學生在實驗過程中遵守了指導,避免了頭部大幅度運動)。
在将上述方法應用于每個資料,在計算個體-小組同步性(TI)時使用EEG殘差,而不是最初的非回歸資料,結果如圖S2E所示。
S2A emotiv系統資料品質評估。
作為emotiv系統資料品質的初步“完整性檢查”,對比睜眼(EO)和閉眼(EC;各2分鐘)EEG。圖S2A右示例學生的閉眼和睜眼枕葉電極記錄的2秒原始EEG。從樣本原始EEG中可以清楚地看到,EC靜息伴随着強α活動而EO靜息期間明顯減弱。圖S2A左顯示了第一個記錄日所有學生的平均功率譜,EO-EC α功率差異明顯。
S2B 雙音調ERP。
學生被動地聽以1000ms SOA交替(音調持續400毫秒;外部揚聲器)播放100個250Hz音調和100個1000Hz音調。在刺激前-200ms到後800ms對EEG資料分段。圖S2B 顯示F4上出現音調典型ERP反應:清晰的N1-P2,250Hz音調(P2峰值)兩峰值略早于1000Hz(P2峰值)音調),結果與成熟裝置報告一緻。
S2C 驗證TI作為共享刺激腦同步測量名額。
為了驗證TI(完全互相依賴)有效性,我們首先計算了每個參與聲調實驗的學生(整合250hz和1000hz刺激)的個體-小組總體互相依賴性。如圖4所示,如果總互相依賴反映了對外部刺激的同步(注意調節作用),那麼TI值應該與單一來源聽覺輸入的朗讀或授課教學方式相當。如圖S2C所示,個體-小組TI值在數值上确實與授課和朗讀教學方式的值相似(隻有記錄一次的5名學生的音調實驗資料,而授課和朗讀資料是多個課時的平均)。
S2D驗證TI作為共同注意測量名額。
從手指移動對照實驗中收集三對被試EEG資料。在基線條件下,兩名被試面對面坐着沒有強制眼神交流。在共同動作環節,随機挑選一名被試為上司者,跟随者跟蹤模仿其手指動作,然後進行角色轉換。基線和共同動作共持續2分鐘。第二個對照實驗比較了同一任務中的合作和競争。在一個區塊設計任務中同時記錄兩個被試EEG。被試必須重新排列色塊以比對圖案。兩個被試要麼互相競争,要麼互相協作。在競争條件下,各被試獨立工作,努力在其他被試之前完成任務。在合作的情況下,兩個被試一起工作以便最短時間内完成任務。 這些發現證明了完全互相依賴是捕捉腦同步性差異的有效方法,與需要協調視覺注意的驗室任務的腦同步增加的發現一緻。
S2E 驗證TI作為共同注意同步神經活動的測量名額。
為了確定同步性未被頭動或殘餘眼動所影響,在對殘餘運動僞影進行回歸分析後,重新分析第5天的資料,原因有兩個:完整的4個教學方式和大多數TI值可用。如圖S2G所示,教學方式之間或教學方式内部的原始資料和重新分析的資料之間沒有一緻性差異。此外個體-小組同步在視訊和讨論方式的差異和其在朗讀-授課方式上的差異幾乎相同。是以,大腦同步性并不是由被試之間的眼睛或頭部同步運動顯著驅動的,且回歸資料時表現為可靠的TI減少。
S2F 在第二所學校複制個體-小組TI。
為了評估在不同時間和背景下的研究發現的穩定性,除了11次重複實驗外,在不同高中重複研究。圖2SF顯示另外一所高中學生視訊或授課的個體-小組TI值(隻包括這兩種教學方式,第一組學生中這兩種風格的資料最幹淨)。與授課相比,視訊方式下個體-小組TI顯著升高與第一組學生的結果相同。
圖S2 對照實驗
A右,示例學生的閉眼和睜眼枕葉電極記錄的2秒原始EEG;A左,所有10名學生的兩個枕部電極在睜眼和閉眼時的平均功率譜。
B從代表性電極(F4)記錄的250Hz(藍色)和1000Hz(紅色)音調事件相關電位。
C音調、教學方式的總互相依賴性。
D來自兩個共同注意控制實驗的資料顯示共同注意作用條件産生的成對TI值始終較高(1-3取自對照實驗1;4-5參與對照實驗2,見正文)。
E在回歸出頭部運動和殘留的眼動僞影(見正文)後,重新計算個體-小組TI值。個體-小組同步性沒有顯著變化。
F另外一所高中學生資料顯示,視訊教學中學生與學生之間的互動程度始終高于課堂授課,與正文結果模式相同。
EEG資料分析:圖S3總結了預處理和分析過程。
腦電預處理:
首先,提取每種教學方式對應的腦電片段。然後對信号進行0.5Hz-35Hz帶通濾波,并将信号分為1秒時間段進行僞影剔除和EEG分析。通過+/-100μV的剔除門檻值來自動剔除具有大運動僞影的資料段(圖S3A)。然後,通過可視化檢查每份資料并手動剔除帶有眨眼和肌肉僞影資料片段(圖S3B)。任何被試某種教學方式下如果超過50%資料段被排除,那麼将該方式下此被試被剔除。每種教學方式(剔除被試前)的平均解釋率見表S1,由于自然環境中無法将被試眼動和身體運動減至最小,是以産生的僞影比例遠遠大于傳統實驗室或基于虛拟現實的實驗中的僞影比例。在去除眼動和其他運動僞影後,超過平均電極振幅四個标準差的電極被排除。
表S1不同教學方式下資料接受百分比,跨被試和記錄日(單個被試資料)。
| 教學方式 | 資料段接受比 | 标準誤 |
|---|---|---|
| 大聲朗讀 | 59% | 2% |
| 視訊 | 69% | 2% |
| 授課 | 58% | 3% |
| 讨論 | 38% | 2% |
計算腦同步性:
完全互相依賴(TI)為了計算每種教學方式下腦同步(圖S3C),采用完全互相依賴(TI)的方法計算給定教學方式下多個學生間的腦同步。使用Welch方法計算的譜相幹性來表征總互相依賴。
對于一對同時獲得的時間序列:(?1, ?1), (?2, ?2),(?3, ?3), … , (?n, ?n),
C(?)是兩個信号?和?在?=?/2?下的相幹性。對于兩個高斯過程,用這個公式測量它們之間的互相資訊總量。TI捕獲了?和?時間序列之間的總線性關系。數值上,對于給定的采樣頻率?s,公式(1)可變為:
Δ?=fs/2(N-1)是頻率分辨率,?是0和Nyquist頻率?/2間隔内所需頻率點數目。
通過計算兩個被試(圖1D和S3E)6個電極一對一Welch相幹度平方來估計TI(完全互相依賴),這兩個被試沒有噪音(也見上文),電極包括兩個枕葉電極(O1,O2)、兩個額葉電極(F3,F4)和兩個頂葉電極(P7,P8)(不包括可能被拒絕的任何電極)。通過用Hanning窗逐漸縮小不重疊1s周期(用0延長到4s),并以0.25Hz分辨率進行傅裡葉變換,計算1-20Hz之間幅度平方相幹。對存在至少60個無僞影共同時間段的配對被試進行分析。在每天每種教學方式中,通過對兩兩被試進行上述預處理後存在的重疊時段來選擇共同時段。例如,對于某個授課記錄,學生1剩餘1-10和12-18時段,學生2剩餘2-13和16-18時段,然後選擇學生1和2的共同時段為2-10、12、13、16-18,然後計算TI,如圖S3D。通過平均所有成對電極TI計算一對被試TI(圖S3E),個體-小組TI通過平均該個體和小組其他人之間所有可能成對組合的TI值得到(圖S3F),然後對這些值在多層線性模型進行統計分析(圖S3G;)。通過将8-12 Hz之内的相幹值相加來計算α相幹性。通過平均學生i所在組所有成對TI值(即?,?,?≠?,其中i和j是每天每種教學方式中可用學生索引;圖S3E),得出學生i的個體-小組TI。使用相同方法計算個體-小組α相幹性(圖S3E-G)。使用最小-最大轉換跨條件和記錄對TI和α相幹值進行标準化。組α相幹性能解釋33%組TI。
圖S3 預處理和分析
A 0.5-35Hz帶通濾波後,連續腦電資料被分成若幹1秒資料段進行僞迹剔除。首先根據+/-100μV門檻值自動剔除運動和眼動僞迹,然後通過視覺化檢查.
B 手動剔除。
C.在11記錄日裡采用四種教學方式(朗讀除外,見正文)向12名學生(分析中僅包括10名學生)展示課堂内容。
D 對每一天和每種教學方式下計算每位學生-其他所有學生TI。分析中隻包括兩個學生都存在資料的時段(共同時期)。
E計算每對電極TI,然後平均。
F對于組TI,平均所有可能TI,每天每種教學方式隻有一個值;對于個體-小組TI,平均個體可用TI值,個體每天每種教學方式隻有一個值。
G多層線性模型(10名學生,為期11天,4種教學方式)統計分析TI。
表S2:因變量的事後Tukey-Kramer檢驗(高亮标記有統計學意義的差異,p<0.05)。
每日學生評分
| 教學風格1 | 教學風格2 | 平均內插補點(1-2) | 标準誤 | P值 |
|---|---|---|---|---|
| 讨論 | 授課 | 1.79 | 0.57 | 0.054 |
| 讨論 | 閱讀 | 3.32 | 0.45 | 0.000 |
| 讨論 | 視訊 | 1.60 | 0.47 | 0.038 |
| 授課 | 閱讀 | 1.53 | 0.55 | 0.089 |
| 授課 | 視訊 | -0.19 | 0.43 | 0.969 |
| 閱讀 | 視訊 | -1.72 | 0.29 | 0.002 |
| 學期後學生評分 | ||||
| 教學風格1 | 教學風格2 | 平均內插補點(1-2) | 标準誤 | P值 |
| 讨論 | 授課 | 3.50 | 0.50 | 0.000 |
| 讨論 | 閱讀 | 4.10 | 0.48 | 0.000 |
| 讨論 | 視訊 | 1.50 | 0.54 | 0.086 |
| 授課 | 閱讀 | 0.60 | 0.31 | 0.269 |
| 授課 | 視訊 | -2.00 | 0.49 | 0.013 |
| 閱讀 | 視訊 | -2.60 | 0.40 | 0.001 |
| 組同步教學風格1 | 教學風格2 | 平均內插補點(1-2) | 标準誤 | P值 |
| 讨論 | 授課 | 0.0112 | 0.0036 | 0.114 |
| 讨論 | 閱讀 | 0.0130 | 0.0060 | 0.278 |
| 讨論 | 視訊 | -0.0040 | 0.0040 | 0.747 |
| 授課 | 閱讀 | 0.0018 | 0.0051 | 0.983 |
| 授課 | 視訊 | -0.0152 | 0.0035 | 0.039 |
| 個體-小組同步教學風格1 | 教學風格2 | 平均內插補點(1-2) | 标準誤 | P值 |
| 讨論 | 授課 | 0.0099 | 0.0026 | 0.019 |
| 讨論 | 閱讀 | 0.0083 | 0.0032 | 0.109 |
| 讨論 | 視訊 | -0.0018 | 0.0032 | 0.940 |
| 授課 | 閱讀 | -0.0016 | 0.0037 | 0.971 |
| 授課 | 視訊 | -0.0117 | 0.0026 | 0.007 |
| 閱讀 | 視訊 | -0.0101 | 0.0047 | 0.203 |
【統計分析】
為了檢驗學生評分(日常評分和學期後評分)和大腦同步性(個體-小組同步性和小組同步性)在兩種教學方式之間是否存在顯着差異,采用重複測量雙向方差分析,其中教學方式和錄制日期是主要因素。對學期後學生評分,僅考慮教學方式。方差分析後進行事後Tukey-Kramer檢驗以評估成對比較的各自貢獻(例如,授課與視訊比較;表S2)。使用日期-個體嵌套多層線性模型(盡可能對所有随機效應進行模組化)計算個體-小組TI和日常問卷得分的相關性。為了分别評估刺激性質(教學方式)和各個因素(如下分析計劃)的獨立貢獻,首先通過将資料重新組織為兩個主要“條件”的二進制因素:教學方式(刺激屬性),包括讨論-視訊組合和朗讀-授課組合(學生對這兩風格評分沒有差異;圖2)。随後對個體-小組TI進行一系列重複測量分析:教學方式vs學期後評分,教學方式vs自我報告的關注度,教學方式vs小組親和力,教學方式vs移情能力。最後,為了評估班級位置對TI的影響,比較了以下三個類别的成對TI值和成對親密度等級:課前眼神交流(相鄰+面對面),課前沒有眼神交流(相鄰+沒有面對面)和不相鄰。
分析計劃:
| 分析 | 動機/研究問題 | |
|---|---|---|
| 刺激與評級的主效應:小組與個人、每日與學期 | ||
| (1)每日和學期後評分的雙向重複測量方差分析 | 計劃 | 學生對教學方式的自我報告是如何因地制宜的? |
| (2)組TI和個體-小組TI的雙向重複測量 | 計劃 | 組矩陣或個體矩陣是否能更好地捕獲資料中的差異? |
| (3)相關性:學生評分x TI | 後續執行 | 學生評分和大腦同步性有關系嗎? |
| 刺激與個體差異對腦同步性的獨立影響:2個“狀态”變量和2個“特質”變量 | ||
| 個體“狀态”差異 | ||
| (1)重複測量多水準線性回歸分析教學方式x評分對個體-小組TI的影響 | 計劃 | 學生對材料的參與是否獨立地預測了個體-小組同步性,而不依賴刺激性質(教學方式)? |
| (2)重複測量多水準線性回歸分析教學方式x關注度對個體-小組TI的影響 | 計劃 | 作為注意力的代表,關注度是否能獨立地預測個體-小組同步性? |
| 個體“特質”差異 | ||
| (1)重複測量多水準線性回歸分析教學方式x群體親和力對個體-小組TI的影響 | 計劃 | 天生喜歡群體的學生會有更大的神經同步性嗎? |
| (2)重複測量多水準線性回歸分析教學方式x移情能力對個體-小組的影響 | 計劃 | 移情能力強的學生是否與組内學生有更大的神經同步性? |
| 共同在場的影響:教師親和力和學生親密度(共同注意與課堂結構) | ||
| 教師親和力 | ||
| (1)教師親和力x個體-小組TI相關性 | 計劃 | 學生對老師的評價是否與更大的神經同步相關? |
| 學生親密度 | ||
| (1)單因素方差分析比較相鄰面對面vs相鄰無面對面vs非相鄰配對TI | 後續執行 | 在課堂上,共同在場(班級結構)或社會互動(面對面)能更好地預測同步性嗎? |
| (2)相鄰面對面學生的親密度x配對TI相關度 | 計劃 | 面對面交流(眼神交流)是否能調節課堂上親密度和同步性之間的關系? |
| (3)相鄰非面對面學生親密度x配對TI相關度 | 後續執行 | 實體鄰近性是否調節課堂上親密性和同步性的相關性? |
| (4)非相鄰學生親密度x配對TI相關度 | 後續執行 | 課堂上親密度和同步性與直接互動或實體接近相關嗎? |
| 注意相關事後分析 | ||
| (1)α相幹性xα功率相關性 | 後續執行 | 這是一項事後分析,提供了額外間接證據支援注意對個體-小組同步性的解釋。 |
【結果和讨論】
本研究關注TI(完全互相依賴)與課堂參與度、社會動态之間的關系。課堂參與度被量化為對不同教學方式評分(圖1B)和學生日常自我報告。根據社交相關人格特質(群體親和力、移情能力)以及課堂互動中社交親密程度(學生與老師之間)量化課堂社會動态。
腦同步與課堂參與
首先研究了大腦同步性(TI)與随時間變化的有關四種不同教學方式學生評分之間的關系。每次記錄後學生對每個部分進行評分且在學期結束後提供每種教學方式的總體評分(圖1A和1B;)。教學方式對兩種學生評分(以教學方式和時間為主因素的重複測量雙向方差分析;日常評分:p <10-5;學期後評分:p <10-8)和腦同步(組同步: p <0.0005;個體與小組同步:p <0.005;)都有顯着的主要影響。總體而言,與聽老師大聲朗讀或授課相比,學生更喜歡觀看視訊和進行小組讨論(圖2A,左圖),這種效果在學期後評分中更為明顯(圖2A,右圖)。小組同步(圖2B,左)以及個體-小組同步(圖2B,右;表S2)的模式非常相似。個體-小組同步性與學生評分之間顯示出很強的正相關:學期後學生評分越高,則時間平均個體-小組同步性越強(p <0.0001;圖2C;圖2A,右顯示相同資料按條件和個體平均分布)。日常評分與小組同步性不相關。
圖2教學方式和個體差異對腦同步性的獨立貢獻
A四種教學方式的每日平均(左)和學期後(右)學生評分:朗讀,視訊,授課和讨論會。誤差線代表标準誤。
B四種教學方式的每日平均組TI(左)和個體-小組TI(右)。誤差線代表标準誤。
C學期後評分雖然對個體-小組同步性具有主要影響,但并未獨立預測教學方式對個體-小組TI的影響。D–F學生關注度(D),小組親和力(E)和移情能力(F)預測了不同教學方式下個體-小組TI。趨勢線按教學方式顯示(藍色:讨論和視訊;黃色:朗讀和授課)。所有值都按0–1範圍(最大-最小)标準化,一個點代表一個學生在四種教學方式下所有記錄日期平均TI。
腦同步純粹是由刺激驅動的嗎?
為了探索刺激屬性(即,教學方式)“獨立”解釋了多少腦同步,個體差異對同步有何貢獻,進行了一系列多元回歸分析以評估TI與許多個體變量(評分,關注度,群體親和力和移情能力傾向)之間的關系,其中包括教學方式作為刺激屬性的一個因素。學期後評分雖然對個體-小組同步性具有主要影響(p <0.0001),但并未獨立預測教學方式對個體-小組TI的影響(學期後評分:p = 0.1327和教學方式:p = 0.1581;圖2C);相反,學生關注度預測了個體-小組同步且與教學方式無關:某天更專注的學生同步性更高(關注度: p = 0.0331和教學方式: p = 0.0004;圖2D)。随後又研究了腦同步與人格特質特别是與群體親和力和共情傾向之間的關系。小組親和力和移情能力都可以獨立于教學方式來預測個體-小組同步(小組親和力:p = 0.0163和教學方式:p = 0.0060;移情能力:p = 0.0185和教學方式:p = 0.0062)。以上結果表明,個體因素(專注和人格特質)在刺激之外促進了腦同步。
腦同步和課堂社會動态
研究結果表明,腦同步是由刺激屬性(教學方式)和個體差異(學生專注度,教學方式偏好,老師喜歡程度和人格特質)共同驅動的。但是,這不能直接說明其他人的存在是否會影響上課期間的同步性。例如,即使在沒有其他人的情況下,共情傾向也會影響大腦相似性。
為了直接解決課堂社會動态問題,收集了學生與老師和其他學生的社會親密度等級,并設定了直接或不直接社互動動的條件。為了研究教師對個體-小組同步性的影響,比較了教師最少參與(視訊)和最大參與(聽課)兩種教學方式。圖2D說明,盡管與整體個體-小組同步性不相同,但視訊條件下同步性始終比聽課好(p = 0.007;表S1)。這種差異與學生對老師的評價相關:學生對老師的評價越好,學生在視訊(老師沒有扮演角色)和聽課(老師扮演不可或缺的角色)之間的同步性差異就越小(圖2E;p = 0.018)。
然後,個體-個體同步性是否随教室中位置(實驗者随機配置設定學生座位)和學生互動(圖1B和3C,學生在課前2分鐘與指定的同伴進行面對面眼神接觸)而變化,即比較以下三種個體-個體同步與自我報告親密程度之間的關系(圖3D):相鄰+面對面,相鄰+沒有面對面,以及不相鄰。與其他兩種情況相比,相鄰面對面學生表現出最高的配對同步性(圖3E; p = 0.0047)。此外,腦同步性與學生互相親密度等級相關,但僅适用于相鄰的+面對面個體:彼此報告較高親密程度的學生在社交活動中表現出較強腦同步性,前提是課前有眼神交流(p = 0.0082;圖3F,實心綠點和實線;該條件對TI -緊密度相關性的影響很小p = 0.0654)。總之,課前面對面交流不僅增加了上課期間的腦同步性,而且似乎可以作為人際關系特征的“激活器”:真實的共同注意而不是被動的共處預測個體-個體腦同步。
圖3 大腦同步性預測教室内社會動态
A、B每名學生視訊vs授課條件下個體-小組TI差異(A)(誤差線代表跨記錄日标準誤)與教師好感度評分負相關(B)(每個點代表一個學生;TI值是所有記錄日平均值;學期結束後學生給出教師好感度評分)。C課前,學生面對面進行2分鐘的眼神交流。D一名學生(綠色圓圈)基線的示意圖,彼此相鄰坐着并在上課前進行眼神交流(相鄰+面對面,adjacent + face-to-face),彼此相鄰沒有眼神交流(相鄰+沒有面對面,adjacent +no face-to-face),以及未相鄰(不相鄰,non-adjacent)。課前面對面有眼神交流的同伴表現出最高的同步性(誤差線反映了學生配對标準誤)。F配對TI與相鄰+面對面對(實心深綠色)的親密關系相關,但與相鄰-不面對面對(實心淺綠色)或不相鄰對(未填充)的親密關系不相關。每個點代表一對學生所有教學方式下平均值。所有值都标準化為0–1範圍(最大-最小)。
共同注意可能是腦同步的來源
腦同步本身并不是一種機制而是對支撐某些心理過程的神經計算的可測量反映。為了更好地了解觀察到的同步效應,需要将諸如注意力、移情能力和親密程度等心理建構分解為與神經度量相聯系的基本心理過程。正如上面已經簡要讨論過的那樣,發現個體-個體同步性與上課期間互相親密程度相關,但僅針對在上課之前進行過眼神交流的學生,這一發現與研究表明眼神交流為共同關注設定了基礎。共同注意(共同的意向性)構成了發展在内的一系列社會心理學社會認知架構,并為需要視覺注意協調的實驗任務中腦同步增加提供了可能的解釋。
研究推測刺激屬性(教學方式)、個體差異(關注度,參與程度和人格特質)以及社會動态(社會親密性和社會互動)均在神經水準上調節了注意力,反過來注意影響個體對各種感覺輸入--老師、視訊或彼此--的神經參與。這與行為證據直接相關,後者表明人們在從事需要共同注意的任務(瞳孔擴張、手勢、行走)時會在身體上(通常是下意識)互相靠近。從更廣泛的意義上說,作為共同注意力的函數個體-小組同步直接取決于一系列電生理結果,這些結果表明當注意參與時鎖定于聽覺和視聽輸入節律的腦節律會增大。
為了提供更多有關共同關注的證據,研究了個體-小組同步性與α功率的關系,α是良好的注意力指數表征。正如預期,學生α活動減少,同時個體-小組α相幹性也随之增加(p = 0.0044)。
這項研究表明,随着“注意力”通過“調節”神經震蕩适應環境變化來調節神經同步,腦同步性會增強。較少參與刺激的人與其他人相比,腦對大腦的同步性較低(圖4),面對面互動的人彼此同步增加。
與實驗室腦電實驗相比,在自然環境下同時記錄一組青少年腦電圖資料無法達到實驗室研究的嚴格水準,但會盡可能采用結構化設計,同時盡量減少學生在正常情況下的互動和課堂内容的參與。其次,11天相同實驗條件下腦電記錄,基本上是對同一組學生重複進行11次相同實驗(圖1A)。最後,通過一系列實驗來驗證可解釋的記錄,并且TI可靠地表征了實驗室和教室環境中個體間神經信号同步性(圖S2)。
圖4 共同注意作為腦同步的一種可能解釋
腦同步的一種可能共同注意示意圖。神經對外部刺激(視訊,老師或彼此的刺激)是由刺激特性(箭頭從“刺激”向下)和注意力(箭頭指向刺激)共同驅動的。在“注意力不足”的情況下,神經振動無法與外部刺激(視訊,老師或彼此刺激)同步(i)。在“共同注意”條件下,α震蕩會減弱并與外部刺激(視訊,老師或彼此吸引)同步(ii)。更專注狀态、具有更多社交參與型人格特質或者直接互動的學生可以調節神經震蕩與刺激(老師,視訊或彼此)同步的程度(iii)。
結論
在一學期11個不同教學日中重複記錄12名學生同時進行課堂活動和社互動動時的大腦活動發現學生間腦同步始終預測着課堂參與度和社會動态。這表明,腦同步性是可以預測教室動态互動的敏感标記,并且這種關系可能是由小組内共同關注引起的。本研究提供了一個有前途的可以研究自然生态環境下群體互相作用神經活動的方法。
原文:Brain-to-Brain Synchrony Tracks Real-World Dynamic Group Interactions in the Classroom